
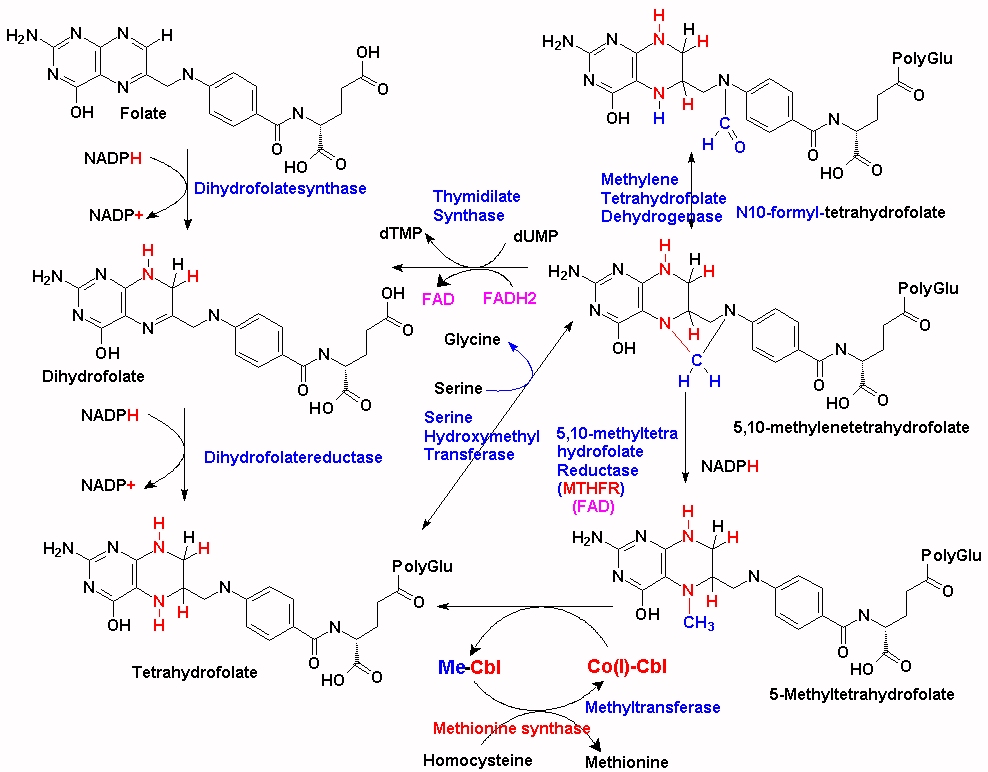
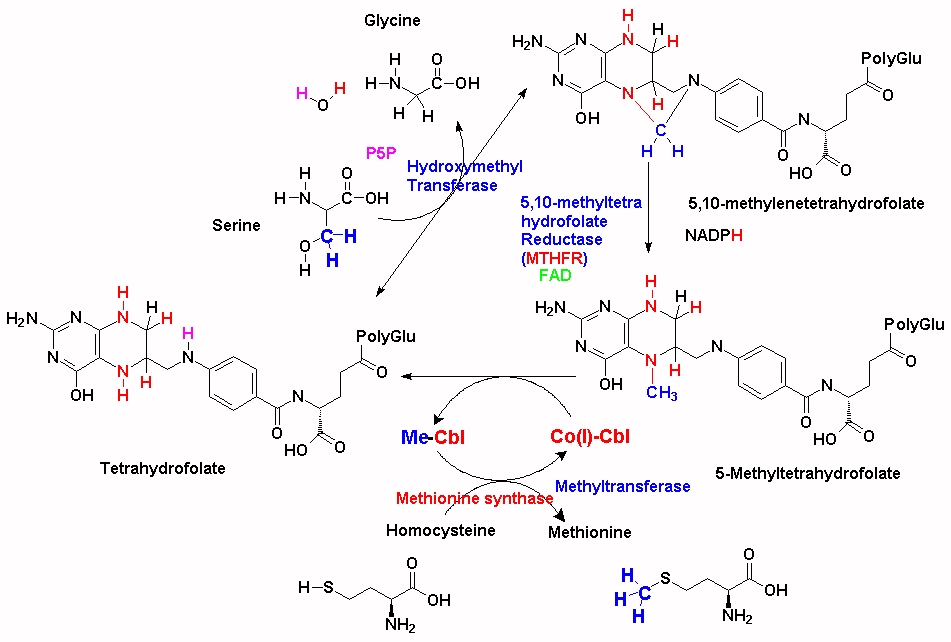
There are over 200 methylation reactions within the cell, the majority of which require S-adenosylmethionine (SAM) as the methyl donor in the reaction. The production of SAM is intimately associated with the uptake of dietary methionine, the folate cycle and methylcobalamin. Within the folate cycle, the Tetrahydrofolate (THF) backbone is substituted with a methylene group by the action of the enzyme Serine-Hydroxymethyl transferase (SHMT) and the product 5,10-methylene-THF is formed. The product, 5, 10 methylenetetrahydrofolate can then be converted to 5-methyltetrahydrydrofolate (5-MTHF) by the enzyme 5,10 methylenetetrahydrofolate reductase (MTHFR). Of note in this reaction is that MTHFR uses FAD (derived from riboflavin, vitamin B2) as an essential cofactor. Without this cofactor the enzyme, MTHFR will not function. In addition, the reduction that occurs also requires input from NADH + H+ (derived from nicotinamide, vitamin B3). Thus, maintenance of functional Vitamin B12, which also requires 5MTHF is critically dependent upon sufficient FAD and NAD. This appears to be a much over-looked fact in the many vitamin B12 supplementation studies that have been carried out, on the treatment of vitamin B12 deficiency in general, and in review after review on vitamin B12 deficiency (Green etal, 2017). The importance of the action of the enzyme can be seen in MTHFR deficiency, which can result in limb weakness, incoordination, paresthesiae, and memory lapses (Haworth et al, 1993), and also in demyelination (Hyland et al, 1988). In the absence of these two vitamins (B2/B3) the enzyme is not functional. In this situation, there is a lack of production of 5MTHF, and a subsequent reduction in the remethylation of Co(I)B12, with subsequent loss of methyl B12 activity and thence lower production of SAM. The rate of reaction of the enzyme is critically dependent upon the concentration of both FAD and NADH. Thus, as the concentration of the two critical co-factors FAD and NADH drops, so too does the rate at which MTHFR converts 5,10-methylene-THF to 5-MTHF.
In the absence of vitamin B12 (Co(I)-Cbl), dietary 5-MTHF cannot enter the folate cycle and is not polyglutaminated, and cannot accumulate inside the cell, with the result that the levels of available intracellular folate are reduced. In the absence of methylcobalamin, the levels of homocysteine, an end product of the methylation cycle, are gradually raised, eventually being excreted from the cell and increasing in serum. Elevated homocysteine is associated with numerous conditions, including an increased risk in cardiovascular disease, regional brain atrophy, mental retardation, seizures, ectopia lentis, secondary glaucoma, optic atrophy, retinal detachment, skeletal abnormalities, osteoporosis, neurological dysfunction, epilepsy and psychiatric symptoms..
.
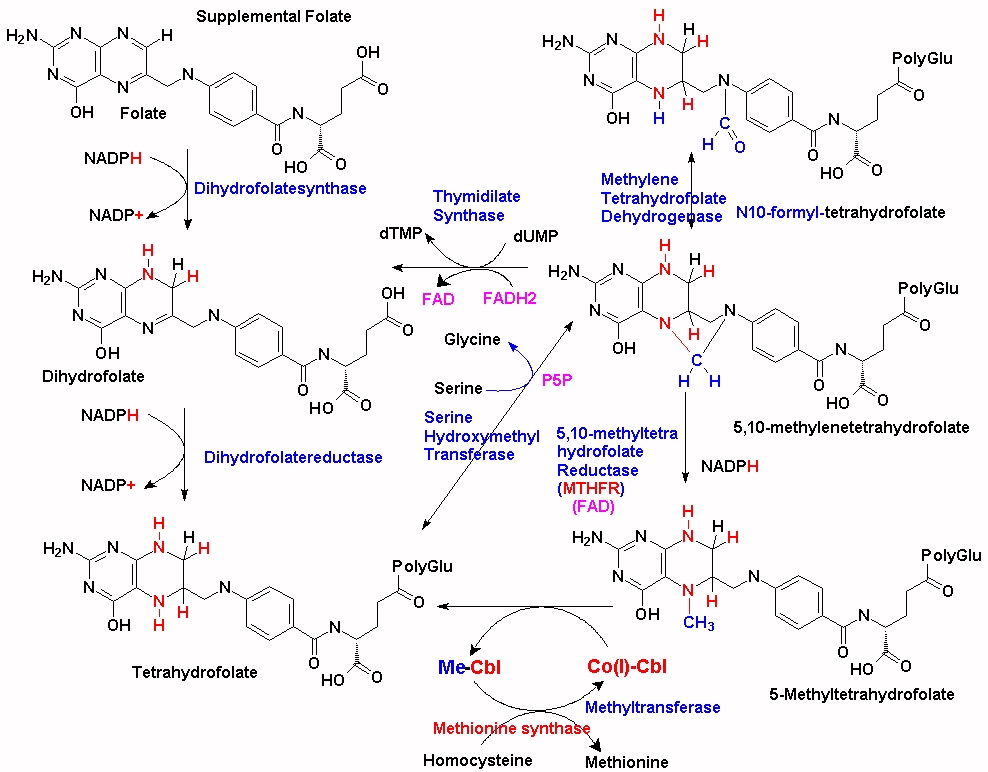
The majority of the methyl groups that are used to make 5MTHF in the cell come from serine, via the enzyme serine hydroxymethyltransferase (SHMT), which forms 5,10-methylenetetrahydrofolate. (See JPG). This is then converted to 5MTHF by MTHFR and then transferred to the methylation cycle as 5MTHF. In order for the methyl group "acquired" during the folate cycling to enter the methylation cycle, vitamin B12 (Co(I)-Cbl) associated with methionine synthase accepts the methyl group from 5MTHF which then forms both methyl-cobalamin (MeCo(III)Cbl) and tetrahydrofolate (THF). THF can then enter the folate cycle. Methionine synthase in turn transfers the methyl group from MeCo(III)Cbl to homocysteine, which is converted to methionine, and Co(I)Cbl is reformed.
Apart from its dependence upon the concentration of both FAD and NADH, the activity of the enzyme can be affected by various mutations in the protein. Mutations that affect function generally involve the binding of FAD to the protein, and are very sensitive to FAD concentration.
There many different mutations in the MTHFR gene, and nearly everyone has one or more, variants of the gene, which may or may not affect the function of the MTHFR enzyme, and hence the rate of conversion of 5,10-methylene-THF to 5-methylTHF. Unfortunately the frequency of such variants is not widely understood in the general population, so much so that a certain amount of web-hysteria has resulted. As an example, one of the common genetic alleles is known as the MTHFRC665T allele (rs1801133). Roughly half the population (or around 3.5 billion people) are heterozygous, CT, for this allele, with 12-20% (depending upon ethnicity) homozygous TT and 30-38% homozygous CC. In persons with some mutations in the MTHFR gene, the enzyme has slightly reduced activity and so the rate at which 5MTHF is produced from the folate cycle is reduced, and so the 5MTHF + Co(I)Cbl <=> THF + MeCbl reaction operates slightly less efficiently. As a result new/incoming supplies of MeCbl must be used for processing homocysteine, leading to a more rapid reduction in vitamin B12. This reduction is most rapid in conditions of low functional vitamin B2 (FAD). The extent of reduction is dependent upon how many and which types of mutations occur in the MTHFR and other methylation associated genes.
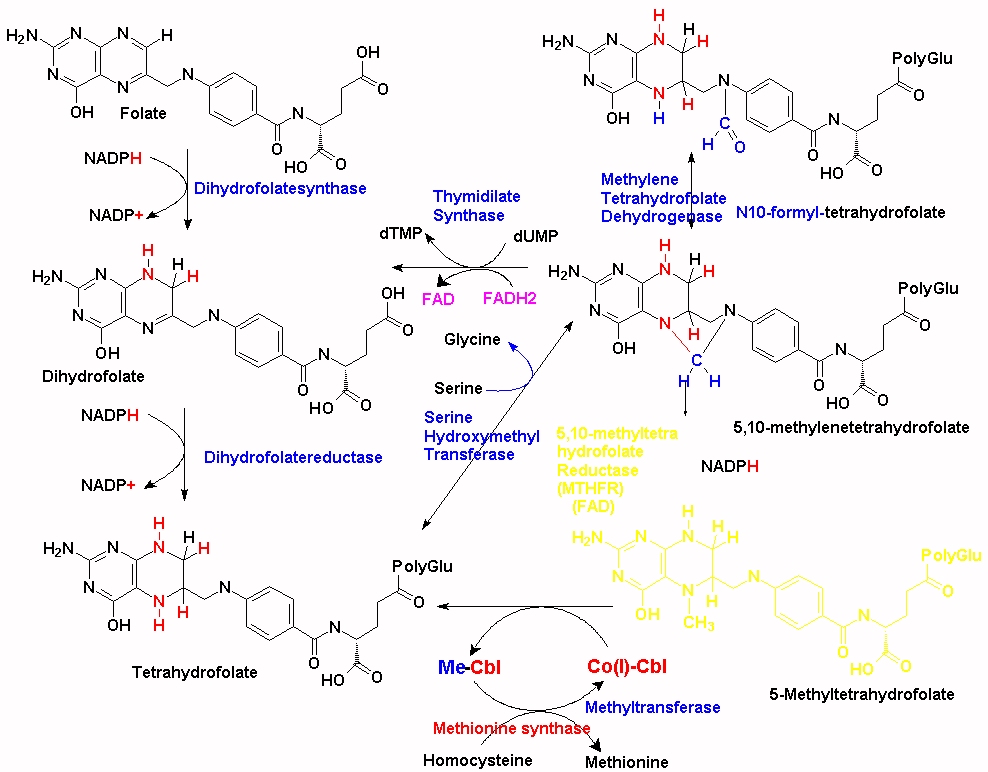
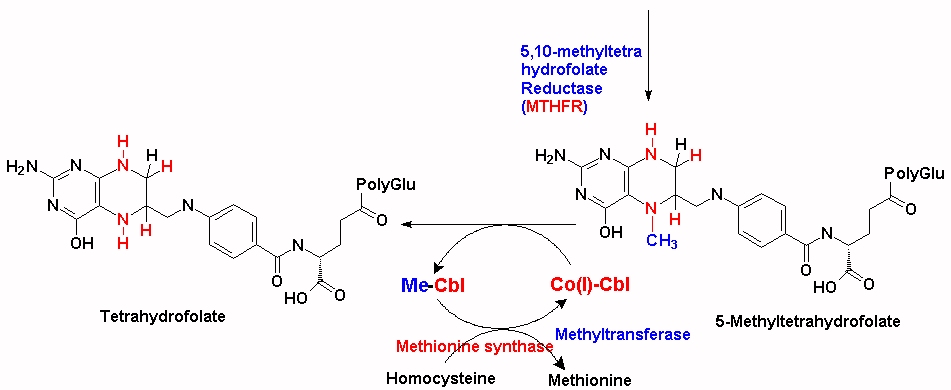
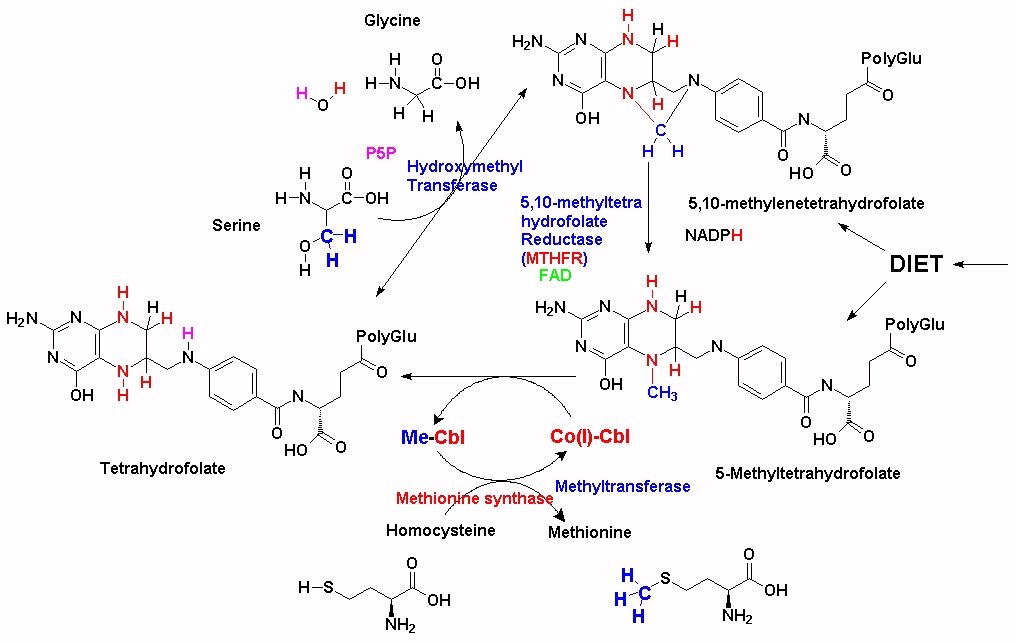
Every methyl group used in synthesis of methionine from homocysteine must be supplied by MeCbl, which can be acquired by supplementation with 5MTHF, but the majority originate via synthesis via SHMT (as outlined above). In the absence of sufficient 5MTHF, there is rapid depletion of VB12 stores. An alternative solution is to supply dietary or supplemental folate as 5MTHF (See Fig2), or increase the amount of folate that is cycling via a folic acid supplement (See Fig3) and Co(I)+Cbl, but as this also is not recycled a new molecule of 5MTHF must always be supplied or the newly synthesized Co(I)-Cbl also will run out. Thus these individuals need a constant supply of both MeCbl AND 5MTHF. An alternative solution is to increase the amount of vitamin B2, which will increase the activity of MTHFR.
Other genetic mutations which affect these reactions are mutations in the genes for methionine synthase, methionine synthase reductase, SHMT and cystathione synthase.
In normal individuals,
MeCbl
reacts with
homocysteine, to
form methionine and
Co(I)Cbl is
formed. If sufficient folate
is present, 5-methyltetrahydrofolate
is produced as an
alternative product of the folate
cycle. In the presence
of
Co(I)Cbl this
methyl group is
transferred from 5MTHF
to
Co(I)Cbl to form
MeCbl
and THF is regenerated.
In vitamin B12 deficient
individuals,
homocysteine
accumulates within the
cell and cannot be used
to convert
5MTHF to THF. This in itself
can lead to functional folate
deficiency within the
cell. Thus, although serum
levels may be normal or
elevated.
Co(I)Cbl is
absent and so the methyl
group from dietary 5MTHF
cannot be transferred
to
MeCbl
and thence to
homocysteine =>
methionine and hence a
deficiency in SAM
results. DNA, RNA and
proteins are not
methylated and this can
lead to defective
function and cancer.
Even in people who start being vitamin B12 sufficient, the constant demand for methylation, and the lack of 5MTHF, leads to the usage of MeCbl and ultimately leads to vitamin B12 deficiency.
Dietary folate is predominantly 5-MTHF, and this must first be processed by methionine synthase +*CoVB12 in order for the folate to enter the folate cycle as THF. In vitamin B12 deficiency this cannot happen and so the folate and the methyl group are "trapped" as 5MTHF and virtually ineffective. Addition of folate (folic acid) as a supplement partially alleviates this as the folate cycle can proceed. One problem with this approach is that methionine is used in protein synthesis and so intracellular methionine stores become depleted. The resultant lack of SAM "turns on" MTHFR, which then pumps the 5,10-methylenetetrahydroflate into 5MTHF synthesis, which, in the absence of VB12 is once again "trapped", thereby consuming the added folate. Addition of folate to MTHFR +/+ individuals reduces the "trapping" of folate, however, residual methionine is lost by further usage in protein synthesis, thus further reducing the intracellular levels of SAM. Supplementation with excess methionine or SAM can boost methylation however it also leads to the increased production of homocysteine, with its own toxicity. It is apparent, therefore, that proper nutritional supplementation would require 5MTHF, plus methyl B12 and possibly methionine (depending upon the nutritional status of the individuals.
Compounding this problem is the need for intracellular folate to be modified with polyglutamate. Without this polyglutamation the folate rapidly diffuses out of the cell. In a curious quirk of fate, 5-MTHF is not a particularly good substrate for the enzyme folylpolyglutamate synthetase, so unless 5MTHF is administered with vitamin B12 and hence allowed to rapidly enter the folate cycle, thereby being polyglutamated, it is rapidly lost from the cell. (see McGuire etal, 1979 http://www.jbc.org/content/255/12/5776.full.pdf )
MTHFR mutations have also been correlated with a higher incidence of various conditions including heart disease, stroke, high blood pressure (hypertension), high blood pressure during pregnancy (preeclampsia), glaucoma, psychiatric disorders (schizophrenia, depression, bipolar disease), and certain types of cancer. The correlation though is somewhat small as too is the increased incidence (generally less than two-fold).
More recently studies have shown a significant correlation between MTHFR TT mutations, reduced vitamin B12 and Parkinson's Disease. A positive correlation has also been shown between MTHFR mutations and rheumatoid arthritis
More recently it has been shown the many people with Chronic Fatigue Syndrome (CFS/ME) have methylation mutations
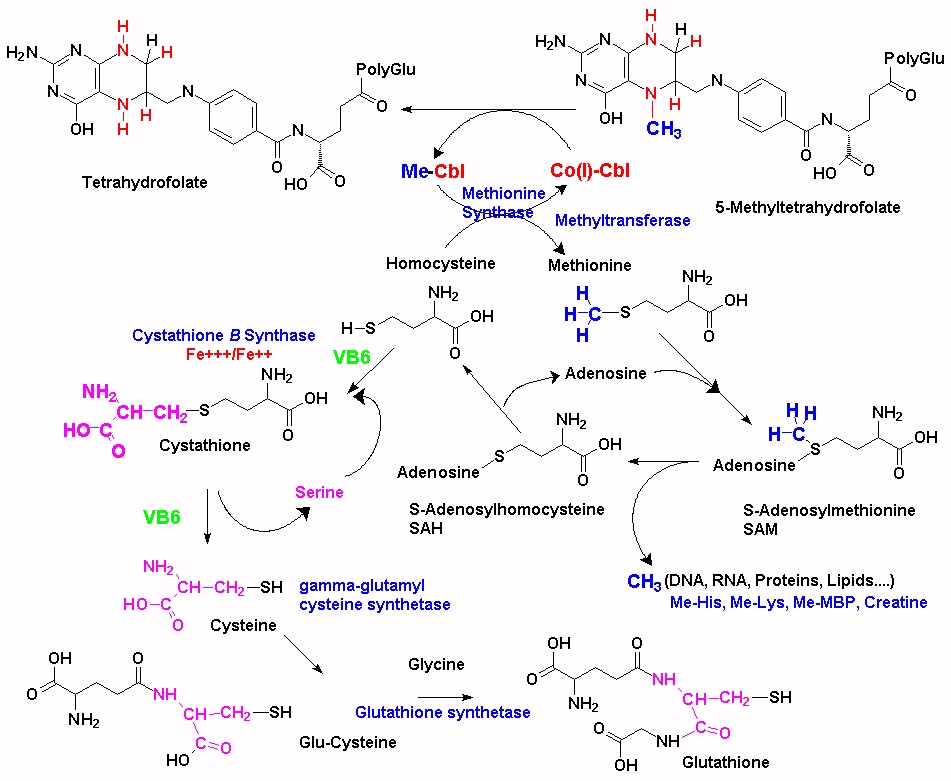
Apart from the formation of methylated myelin basic protein, the are over 200 methylation reactions in the body including the production of creatine (involved in energy production), CoQ10 (an essential electron shuffle vector in the electron transport chain), carnitine (involved in uptake of fatty acids into mitochondria), the production of phosphatidyl-choline, adrenalin, melatonin (essential for neuronal development, gut health, and sleep),myelin basic protein, methylation of lysine, histidine and arginine, and in the inactivation of histamine, dopamine, nor-adrenalin, and adrenalin, amongst others.
Essential in the formation of myelin sheath is its association with myelin basic protein (MBP). In order for MBP to associate with the myelin it must first be post-transationally modified by methylation of arginine. If the arginine is not methylated, the structure of MBP is changed and otherwise buried arginine residues are exposed. These can then act as substrates for nitric oxide synthase (NOS), which converts arginine to citrulline. This then is a different MBP from normal and hence can generate an immune response to MBP and MS may develop. MTHFR mutations have been correlated with MS, as too has VB12 deficiency.
Many people with methylation mutations such as MTHFR have trouble getting pregnant or maintaining their pregnancy. The baby born to an MTHFR positive mother and/or father can inherit the condition. A lack of folate/and or vitamin B12 particularly in these individuals can lead to Spina bifida, anenceaphaly, and Down's syndrome. Elevated homocysteine can lead to low birth weight and premature babies. Studies have now shown that more than 80% of babies with autism have some kind of methylation mutation.
References
Green etal. Vitamin B12 deficiency. Nature Reviews 2017 3: article 17040
Haworth etal, Symptomatic and asymptomatic methylenetetrahydrofolate reductase deficiency in two adult brothers. Am J.Med.Genet, 45, 572-6.
Hyland, etal, 1988 Demyelination and decreased S-adenosylmethionine in 5,10-methylenetetrahydrofolate reductase deficiency. Neurol, 39,459-62
Copyright © 2018
vitaminb12deficiency.info. All Rights Reserved.
The statements on this site compose a compendium of generally recognized signs
of vitamin B12 deficiency, and problems that can then ensue They also are formulated from a summary of relevant
scientific publications. In addition they may contain some forward looking
statements of a general nature.
Reproduction in whole or in part in any form or medium without express written
permission is prohibited
{kind=link}
{kind=link}
{kind=link}